






Bio-Share丨双特异性抗体:结构设计与疾病应用的新前沿
发布时间: May 26 , 2026
目前研究表明,双抗的分子架构直接影响其结合力、稳定性、体内半衰期以及免疫效应功能,这意味着在结构设计时不仅要考虑靶点选择,还必须权衡结构对药效学和安全性的潜在影响[1]。
1、条件激活型双抗的兴起
从适应症分布来看,双抗的主战场依然是肿瘤领域,占比高达86%。其中,76%用于实体瘤,24%用于血液恶性肿瘤。目前,双抗的成功案例仍然主要集中在血液瘤。在T细胞衔接器中,57%用于实体瘤,43%用于血液恶性肿瘤,然而实际获批上市的分子绝大多数仍是针对血液瘤的产品。
究其原因,这两类疾病在生物学属性上存在根本性差异。血液瘤的肿瘤细胞悬浮于血液和骨髓中,T细胞可以相对直接地接触肿瘤细胞,肿瘤抗原的表达也相对均一。而在实体瘤中,肿瘤微环境的复杂性——基质屏障、缺氧、肿瘤抗原异质性、免疫抑制性细胞浸润等,构成了多层次的障碍。随着临床数据的积累,研究人员的主要问题变为:双抗能否在正确的位置、以足够的局部浓度、在足够的持续时间内发挥功能?
近年来,条件激活型双抗(conditionally active antibodies)的出现提供了一种新的解决方案。这类分子被设计为在正常生理条件下处于“沉默”状态,只有当进入肿瘤微环境后,在特定的局部信号(如:肿瘤相关蛋白酶切割)作用下才被激活。其结构为:在循环系统中保持沉默以避免对正常组织的损伤,在肿瘤局部被“唤醒”以发挥治疗作用。这种结构设计使得双抗可以在保留全身安全性的同时,实现更强的肿瘤局部效应。
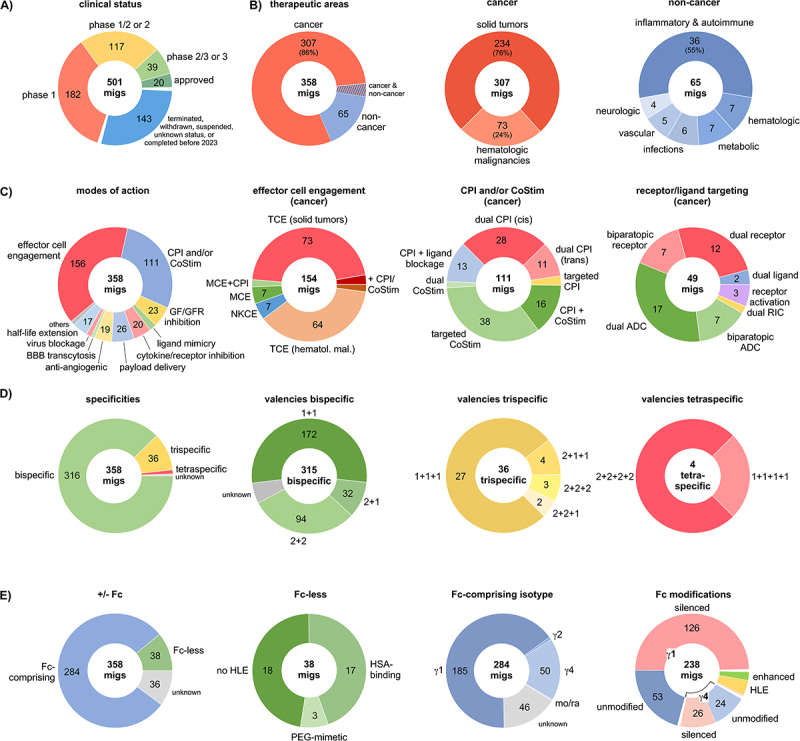
图1:多特异性免疫球蛋白在临床应用或研发中的特性。来源:Mabs 2026 Jan 15;18(1):2613548. doi: 10.1080/19420862.2026.2613548
2、不同结构双抗的临床逻辑
在已确认结构形式的多特异性免疫球蛋白(migs)中,最常见的类别是T细胞衔接器(TCEs),占44%;免疫调节类(即双重免疫检查点抑制或共刺激激活)占31%。在这些数字背后,每一种结合方式,被认定为真正决定双抗性能的关键信息。
- 效应细胞衔接器:最典型的T细胞衔接器同时结合肿瘤相关抗原(TAA)和T细胞表面的CD3,将T细胞“桥接”到肿瘤细胞附近并激活杀伤功能[2];
- 免疫调节型双抗:同时阻断PD-1/CTLA-4或PD-1/LAG-3等多个免疫检查点,或将检查点抑制与共刺激信号(如4-1BB、CD28)整合于同一分子中[3];
- 双通路阻断型:如EGFR×MET,其价值不仅在于同时阻断两条通路,更在于通过抑制MET通路来阻断EGFR的旁路激活,从而改变耐药的分子机制[4]。
即使在相同的靶点组合(如:CD19×CD3)下,不同结构形式设计的双抗,其免疫突触的几何构型、受体聚集方式、突触稳定时间都存在差异,直接影响T细胞的活化强度、连续杀伤效率、细胞因子释放综合征风险和T细胞耗竭[5]。因此,形式本身正在成为药理学的一部分,逐渐走向疾病应用。
3、双抗在疾病应用的扩展
除了上述强调的肿瘤领域,双抗在疾病方面的应用边界正在被不断拓宽。根据市场,标志性的案例是Emicizumab(FIXa×FX)——它既不是阻断某个靶点,也不是激活某种免疫细胞,而是通过桥接凝血因子FIXa与FX,使FIXa能催化FX活化,从而模拟缺失的凝血因子VIIIa(FVIIIa)的辅因子能力,在凝血系统中重建血友病A患者缺失的生理功能[6]。
这一案例的意义在于:双抗在疾病应用领域的功能正在被重新定义。它不仅可以杀伤肿瘤、阻断免疫检查点、激活免疫细胞,还可以重建生物过程、模拟天然复合体、替代缺失功能、重构信号关系[7]。在某些慢病领域,双抗展现出比ADC更强的适用潜力。
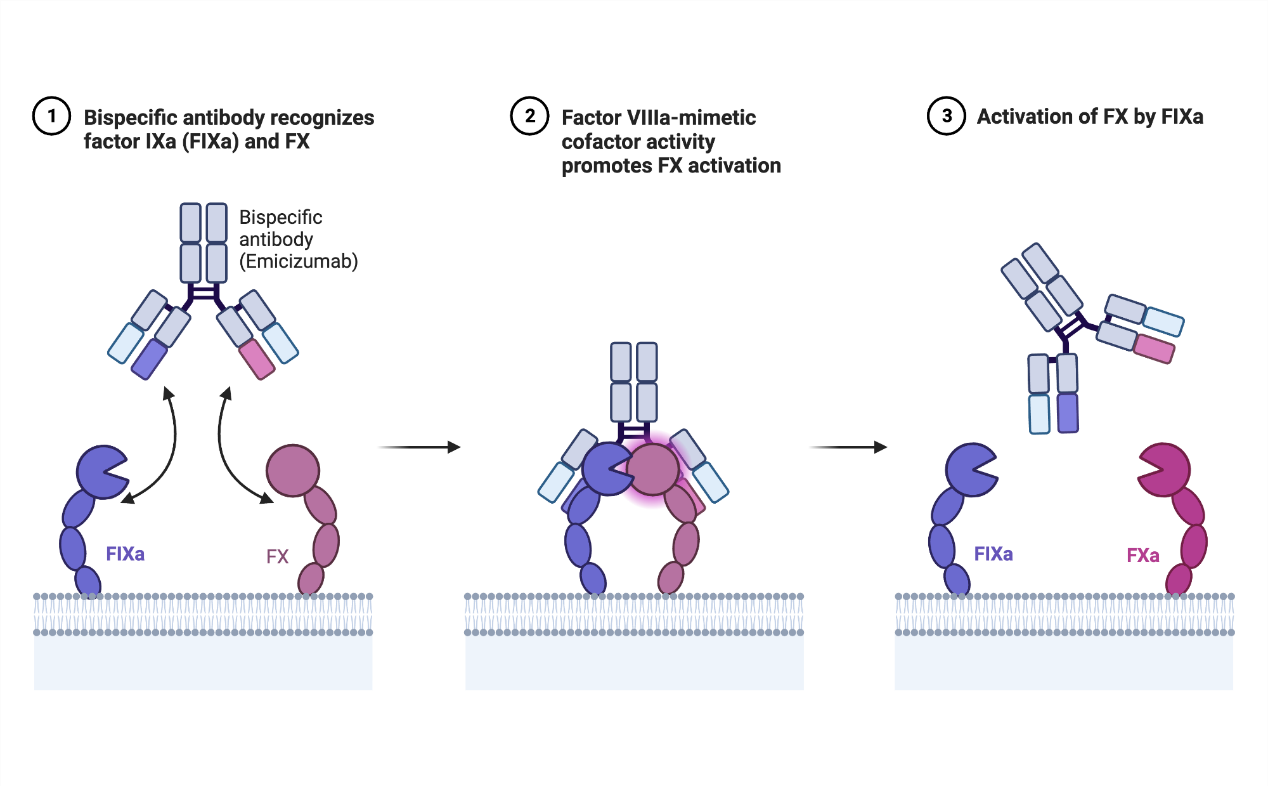
图2:Emicizumab的作用机制。来源:BioRender
总结
双特异性抗体的结构设计,正在从“选对靶点”走向“设计功能”。不同的分子构型,最终影响的是稳定性、免疫突触的有效形成、药物在体内的分布、以及安全性的优良。在肿瘤领域,条件激活型双抗的出现,给实体瘤微环境提供了一个切实可行的思路。从机制上看,T细胞衔接器、免疫调节和双通路阻断这几类设计,已经领跑,而构型本身,也越来越直接地决定了药理学效果。而肿瘤之外,Emicizumab这类分子证明了双抗能做的事远不止杀伤肿瘤——重建凝血过程、模拟天然复合体、替代缺失的生理功能,这些都已经成为现实。未来双抗在更多疾病领域里的应用,值得持续关注。
参考资料:
[1] Brinkmann U, Kontermann RE. The making of multispecific immunoglobulins - a clinical perspective. MAbs. 2026 Dec;18(1):2613548. doi: 10.1080/19420862.2026.2613548. Epub 2026 Jan 15. PMID: 41542910; PMCID: PMC12818813.
[2] Dettling DE, Kwok E, Quach L, Datt A, Degenhardt JD, Panchal A, Seto P, Krakow JL, Wall R, Hillier BJ, et al. Regression of EGFR positive established solid tumors in mice with the conditionally active T cell engager TAK-186. J Immunother Cancer. 2022. June. 10(6):e004336. doi: 10.1136/jitc-2021-004336.
[3] Pang X, Huang Z, Zhong T, Zhang P, Wang ZM, Xia M. Li B.Cadonilimab, a tetravalent PD-1/CTLA-4 bispecific antibody with trans-binding and enhanced target binding avidity. MAbs. 2023. Jan. 15(1):2180794. doi: 10.1080/19420862.2023.2180794.
[4] Neijssen J, Cardoso RMF, Chevalier KM, Wiegman L, Valerius T, Anderson GM, Moores SL, Schuurman J, Parren PWHI, Strohl WR. Chiu ML discovery of amivantamab (JNJ-61186372), a bispecific antibody targeting EGFR and MET. J Biol Chem. 2021. Jan. 296:100641. doi: 10.1016/j.jbc.2021.100641.
[5] Gedeon PC, Schaller TH, Chitneni SK, Choi BD, Kuan CT, Suryadevara CM, Snyder DJ, Schmittling RJ, Szafranski SE, Cui X, et al. A rationally designed fully human EGFRvIII: CD3-targeted bispecific antibody redirects human T cells to treat patient-derived intracerebral malignant glioma. Clin Cancer Res. 2018. Aug 1. 24(15):3611–3631. doi: 10.1158/1078-0432.CCR-17-0126.
[6] Kitazawa T, Shima M. Emicizumab, a humanized bispecific antibody to coagulation factors IXa and X with a factor VIIIa-cofactor activity. Int J Hematol. 2020;111(1):20–30. doi: 10.1007/s12185-018-2545-9.
[7] Beishenaliev A, Loke YL, Goh SJ, Geo HN, Mugila M, Misran M, Chung LY, Kiew LV, Roffler S, Teo YY. Bispecific antibodies for targeted delivery of anti-cancer therapeutic agents: a review. J Control Release. 2023. Jul. 359:268–286. doi: 10.1016/j.jconrel.2023.05.032.







